1980年发现成纤维细胞经Poly I-C刺激后能产生一种抑制病毒复制的细胞因子,称为β2干扰素(IFN-β2)。以后的研究结果未能证实这种因子的直接抗病毒作用,但具有其它多方面的生物学功能,根据实验系统和功能的不同,不被命名为杂交瘤/浆细胞瘤生长因子(hybri-doma/plasmacytoma growthfactor,HPGF),B细胞分化因子(B cell differentiationfactor,BCDF),B细胞刺激因子-2(b cellstimulatory factor 2,BSF-2),26kDa,溶细胞性T细胞分化因子(cytolytic T cell differentiation factor,CDF)和肝细胞刺激因子(hepatocyte stimu-lating factor,HSF)等。1986年统一命名白细胞介素6(interleukin 6,IL-6)。
1.IL-6的产生 淋巴样和某些非淋巴样细胞均可产生IL-6。
(1)T细胞:T细胞产生IL-6依赖于巨噬细胞或PMA。抗原提呈细胞刺激相应的T细胞克隆,以及HTLV-I感染的T细胞系等均可分泌IL-6。
(2)B细胞:如SAC刺激而活化的B细胞。
(3)单核细胞:LPS刺激单核细胞产生IL-6,某些单核细胞系如P388D1也可分泌IL-6。
(4)成纤维细胞:可自发产生IL-6,其它因子或刺激物如IL-1、TNF、PDGF、IFN-β、PolyI-C、A23187、PMA等可促进IL-6的产生。
(5)肾小球系膜细胞、角朊细胞、内皮细胞等在一定培养条件下均可产生IL-6。此外,肿瘤细胞或细胞系如MG63成骨肉瘤,T24膀胱癌、A549肺癌、7860肾癌、SK-MG-4神经胶质母细胞瘤、U373星状细胞瘤、心脏粘液瘤细胞和骨髓瘤细胞等也能分泌IL-6。最近发现垂体前叶中的滤泡—星状细胞(folliculostellate)可产生IL-6,可能与败血症时LPS刺激导致GH、ACTH等激素水平升高有关。
IL-1、TNF、IFN-β、PDGF、LPS、Poly Ⅰ-C、A23187和PMA等对IL-6的产生具有正调节作用。
2.IL-6的分子结构和基因 1985年Kishimoto等从人T细胞中首先获得IL-6cDNA克隆成功,人IL-6基因与小鼠有65%同源性。人IL-6基因位于第7号染色体,长约5kb,有5个外显子和4个内含子。
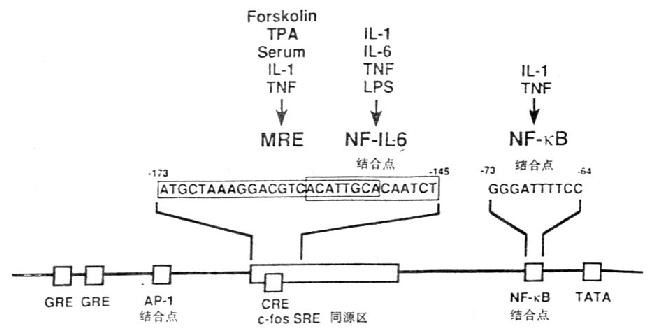
图4-1 IL-6基因的功能调节区
在IL-6基因功能调节区基因中存在着儿种转录控制元件(transcriptional control element),如糖皮质激素反应元件(glucocorticoidresponsive elements,GRE)、AP-1结合位点、c-fos血清反应元件同源物(c-fos serum responsiveelement homology,c-fos SRE homology),cAMP反应元件(cycli AMp responsive element,CRE)和NF-κB结合位点。IL-1、TNF等细胞因子可使IL-6启动子很快发生一过性的活化。IL-1反应的元件在IL-6启动基中-180/-123;IL-6核因子(NF-IL-6)识别一段特殊的14bp,ACATTGCACAATCT。多反应元件(multi-responseelement,MRE)位于c-fos SRE同源区内,这个区域对IL-1、TNF、forskolin和PMA诱导IL-6产生有关;与IL-1、TNF刺激IL-6产生有关的NF-κB位于TATA盒的上游。
人IL-6分子由212个氨基酸残基组成,包括28个氨基酸残基的信号序列,成熟IL-6为184氨基酸残基,分子量26kDa。IL-6分子由4个α螺旋和C端(175~181位氨基酸)受体结合点所组成,其中179位精氨酸残基对于与受体的结合非常重要。分子中糖基对生物学活性功能并非必需,N端23个氨基酸残基虽不直接与IL-6生物学活性有关,但对整个IL-6分子组成起稳定作用。人IL-6氨基酸序列与小鼠IL-6有42%同源性,人的IL-6对小鼠某些细胞有刺激作用。IL-6与G-CSF和IFN-β有较高同源性,对骨髓造血细胞和髓样白血病细胞的某些作用也有相似之处。
3.IL-6的受体 目前已知,IL-6R至少由称之为IL-6结合受体蛋白(IL-6binding receptor protein)和称为信号转导蛋白(signal-transducingprotein)的gp130所组成,习惯上前者称之IL-6R。
(1)IL-6R(CD126):人IL-6R由468个氨基酸组成,切除N端19个氨基酸残基后的成熟分子有449氨基酸,胞膜外区、穿膜区和胞浆区分别为339、28和82个氨基酸,分子量为80kDa,6个N糖基化位点。胞膜外由一个Ig样区(C2,约100氨基酸)、2个Ⅲ型纤维结合蛋白结构(各含100氨基酸)及1个细胞因子受体的同源区所组成,后者含4个保守的Cys和一个WSXWS结构。单独IL-6R与IL-6结合为低亲和力。IL-6R分布于淋巴样细胞和非淋巴样细胞,如活化B细胞、EBV转化B细胞、急性淋巴母细胞白血病细胞、骨髓瘤细胞、静止T细胞、肝细胞、单核细胞、急性髓样白血病(AML)细胞、嗜铬细胞瘤细胞等。
(2)gp130(CDw130):分子量为130kDa的糖蛋白,共有14个潜在N-糖基化位点,胞膜外区、穿膜区和胞浆区分别有597、22和277个氨基酸。胞膜外区有1个IgC2区,6个Ⅲ型纤维结合蛋白的结构,其中第二个和第三个结构区之间有4个保守的Cys和WSXWS结构的区域,形成1个细胞因子受体家族结构特征的结构域。gp130不能直接与配基IL-6结合,在生理情况下,IL-6与IL-6受体结合后使IL-6R的构象发生变化并迅速与两个gp130分子结合,形成高亲和力的结合位点,并通过gp130亚单位传递信号。人和小鼠gp130在氨基酸水平上有77%的同源性。转染gp130cDNA小鼠pro-B细胞在IL-6/sIL-6R复合物刺激下可传递增殖信号。小鼠体内注射IL-6可增加gp130mRNA的表达。目前已证实,gp130除组成IL-6高亲和力受体外,也是白血病抑制因子(LIF)、抑瘤素M(OSM)、睫状神经营养因子(CNTF)和IL-11等受体所共用的亚单位。
(3)信号转导:gp130与IL-6/IL-6R复合物结合后,刺激gp130胞内部分发生酪氨酸磷酸化,目前关于参与此过程的酪氨酸蛋白激酶的作用还不清楚。酷氨酸激酶被激活后继而引起丝氨酸/苏氨酸激酶如丝裂原活化的蛋白激酶(mitogen activated proteinkinase,MAPK)的激活,使NF-IL-6中丝氨酸和苏氨酸磷酸化而被激活,从而促进相应基因的活化。
(4)sIL-6R:存在于正常人尿、骨髓瘤细胞系U266培养上清,PHA活化人PBMC以及HTLV-I阳性细胞也能分泌sIL-6R,分子量为50kDa。用反转录PCR从正常人细胞和骨髓瘤细胞中均分离出编码sIL-6r mRNA,序列分析表明与膜结合受体相应区域序列一致。sIL-6也可从膜结合的sIL-6R(mIL-6)脱落而来。sIL-6R与IL-12p40亚基具有高度同源性,而IL-6与IL-12的p35亚基序列高度同源。因此可以推测类似于IL-6/sIL-6R复合物的IL-12分子可能也通过类似于pg130分子作用于细胞。与其它可溶性细胞因子受体不同,sIL-6R结合IL-6后可与细胞膜表面gp130结合,增强IL-6的刺激活性。而可溶性gp130(sgp130)可抑制sIL-6R/IL-6复合物的活性。sIL-6R水平的升高与某些自身免疫性疾病有关。
4.IL-6的生物学活性
(1)刺激细胞生长:IL-6可促进多种细胞的增殖,如B淋巴细胞杂交瘤、浆细胞瘤、EBV转化的B细胞、T细胞、PMA和IL-4刺激的胸腺细胞、造血干细胞、角朊细胞和肾小球系膜细胞。
(2)促进细胞分化:如B细胞分化和Ig的分泌,CTL分化,协同IL-2增强CTL中穿孔素基因的表达,并增加T细胞IL-2产生和IL-2R表达,诱导异巨噬细胞、神经细胞和NK细胞分化。协同IL-3促进干细胞分化和巨核细胞的成熟。明显促进小鼠骨髓移植后免疫功能的重建。
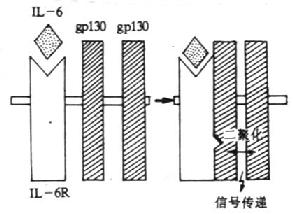
图4-2 IL-6受体信号传递的模式图
(3)加速肝细胞急性期蛋白(acute phase protein)的合成。
(4)抑制M1髓样白血病细胞系的生长,促进其成熟和分化;抑制黑素瘤、乳腺癌细胞生长。
5.IL-6与临床 IL-6与临床上多种疾病的发生有一定的关系。
(1)IL-6与自身免疫性疾病
①心脏粘液廇:患者往往表现为高丙球蛋白血症,有多种血身抗体以及急性期蛋白升高。培养的粘液瘤细胞含有IL-6和IL-6mRNA,患者血清中IL-6明显升高,骨髓中可见有IL-6依赖的多克隆浆细胞增殖。手术后上述症状和体症可见逐渐消退。
②Castleman氏病:患者表现为高丙球蛋白血症,急性期蛋白和血小板升高,增生的淋巴结生发中心中B淋巴样细胞产生IL-6,其中某些患者可发展成为多发性骨髓瘤。
③类风湿性关节炎:表现为多克隆性浆细胞增多症,自身抗体、C反应蛋白(CRP)和血小板升高。急性期血清以及关节的滑液中能测到IL-6,滑液中IL-6与IgG以及血清中IL-6与C反应蛋白之间有明显的相关性。患者的T细胞、B细胞、滑膜细胞以及软骨细胞均可产生IL-6。
④艾滋病:与艾滋病患者多克隆B细胞活化有关,HIV感染诱导单核细胞产生IL-6可能引起血清中IL-6水平的升高。此外,IL-6可能是Koposi氏肉瘤的主要生长因子之一,反义IL-6基因在体内可抑制Koposi氏肉瘤细胞的生长。
(2)IL-6与肿瘤
①浆细胞瘤形成:慢性炎症诱导IL-6生物合成增加与浆细胞瘤的发生有关。如用石腊或降植烷腹腔刺激小鼠,诱导炎症,可诱导出较高比例的浆细胞瘤。在患者也可见到类似情况,如早先发生的类风湿性关节炎可能与浆细胞瘤形成有关。在体外,IL-6可促进浆细胞瘤和骨髓瘤细胞的生长,某些浆细胞瘤细胞生长依赖于IL-6的存在。
②可能通过自分泌机理与非Hodgkin氏淋巴瘤、慢性淋巴细胞白血病和急性髓样白血病的发病有关。血清中IL-6水平与多发性骨髓瘤如浆细胞白血病病情严重程度有关。多发性骨髓瘤患者不仅骨髓细胞表面IL-6R表达增加,而且血浆中sIL-6R水平明显升高。
(3)IL-6与膜增生性肾小球肾炎(mesangial proliferativeglomerulonephritis,MPG):MPG患者尿中可测出IL-6,而且其水平与疾病的发展有关。体外培养的患者肾小球膜细胞可产生IL-6,提示MPG发生与IL-6的自分泌有关。
IL-6转基因小鼠可出现某些与临床上相似的变化,如血清中高浓度的IL-6和IgG1,浆细胞增多症,MPG的发生以及骨髓中巨核细胞的成熟。
(4)烧伤和术后伴有血清IL-6水平增加。
(5)病毒性脑膜炎和脊髓膜炎小鼠的胶质细胞可分泌大量IL-6,IL-6又可促进脑胶质细胞分泌其它神经营养因子和神经生长因子。
(6)IL-6作用于下丘脑-垂体-肾上腺轴,刺激ACTH和皮质激素的释放以及星状细胞合成内啡肽。
IL-1和TNF-α对IL-6引起的病理损伤可能有协同作用。
应用IL-6治疗放疗、化疗所致血小板减少症、癌症以及作为疫苗佐剂已进入临床试验。